Adenylate-forming enzymes
We have a long-standing interest in a family of Adenylate-Forming enzymes that catalyze two step reactions. In the first reaction, a carboxyate group reacts with ATP to form an acyl adenylate and inorganic pyrophosphate. This activated carboxylate then reacts in a second partial reaction. In most enzymes, the subsequent reaction is with a free thiol to form a thioester after the displacement of AMP. However, other partial reactions are known, including amide formation or an oxidative decarboxylation.
Members of this family can be divided into three subfamilies. Acyl- and Aryl-CoA Synthetases. These enzymes ligate carboxylic acids to the thiol of Coenzyme A. The second subfamily are the adenylation domains of the modular NRPS proteins. The third subfamily are beetle luciferase enzymes that catalyze a decarboxylation reaction that leads to an activated luciferin molecule that breaks down to give light.
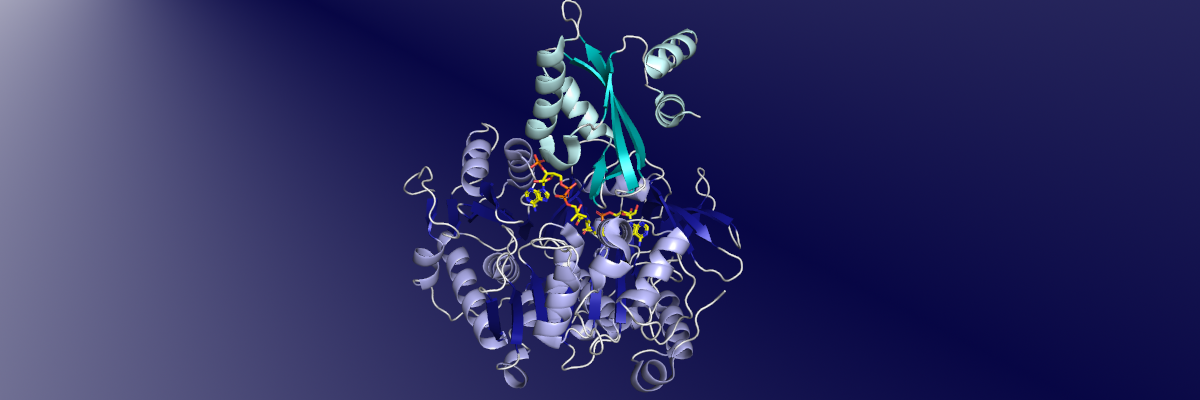
Structural Biology of ANL enzymes
Studies with multiple ANL enzymes from all three subfamilies have demonstrated that the enzymes share common biochemical and structural features. The structures of these enzymes illustrate two subdomains, a larger N-terminal domain of 400-500 residues and a C-terminal subdomain of ~120 residue. These domains have also been called the Acore and Asub domains. The active site for binding the carboxylate substrate and ATP is located between the two subdomains. Structural studies, supported by biochemical analysis, have shown that the enzymes adopt two conformations to catalyze the two partial reactions. Upon completion of the adenylation step, the C-terminal subdomain rotates by ~140° to adopt a second conformation that is used to catalyze the second partial reaction. This conformational change is particularly interesting for the modular NRPS enzymes where this large rotation is used to support delivery of the downstream carrier domain to the active site for thioesterification.
Structures of multiple ANL enzymes have been determined in distinct catalytic states that support this